Scientists recently suggested that the Earth’s sixth mass extinction has begun. As terrifying as that sounds, surely humans are too smart and too important to get wiped out? Paleontologists have long tried to shed light on this question by looking for general rules that might predict the survival of a species.

Scientists recently suggested that the Earth’s sixth mass extinction has begun. As terrifying as that sounds, surely humans are too smart and too important to get wiped out? Paleontologists have long tried to shed light on this question by looking for general rules that might predict the survival of a species.
While this is not exactly a straightforward exercise, research so far indicates that the odds are not in our favor.
Join 500,000+ readers who start their day with Quartz.
By subscribing, you agree to our Terms of Service and Privacy Policy.
Life on Earth can be traced back to a single unicellular species, perhaps some 3.5 billion years ago. Since then, diversity and maximum complexity has increased and millions of species have evolved.
But how did we go from one species to millions of species? Let’s do a simple thought experiment. Lineages can split in two so that one species yields two, two yield four, four yield eight and so forth. If plotting this process as a curve, the number of species would grow exponentially over time. Of course, species will also go extinct, but provided this happens less often than new ones arise, you will still end up with an exponentially increasing curve.
But can diversity go on increasing forever? Charles Darwin certainly thought not, and believed that the Earth probably had a carrying capacity. He likened species to wedges driven into a log, each occupying their own niche or patch of ecospace. As the number of wedges approaches the carrying capacity, it becomes more difficult to insert new ones, until adding new wedges forces older ones out.
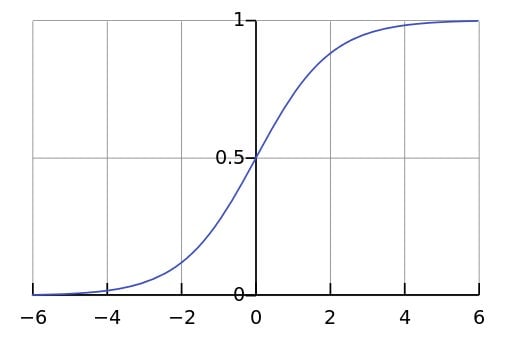
The idea that the Earth can only accommodate a finite number of species modifies our simple model somewhat. Early on in the process, numbers are far from carrying capacity, and growth is exponential. Later on, progressively harder brakes are put on, and the rate of growth slows down, so that diversity reaches a plateau. Together, these forces yield an S-shaped or sigmoidal curve.
So what do we see when we look at the real history of life in the fossil record? Fortunately, paleontologists have systematically compiled catalogs of fossil genera, making it possible to compare. What they show, however, is a much more complex picture.
Some of the earliest diversity curves were produced for marine organisms. These revealed five mass extinction events over the last half billion years, in which diversity markedly and rapidly reduced. The first two of these—the end of the Ordovician, about 444 million years ago, and and the end of the Devonian, about 359 million years ago, occurred at a time when diversity appeared to have reached a plateau. Diversity simply bounced back to previous levels after they struck.
The third mass extinction, dubbed the “Great Dying”, some 252 million years ago at the boundary between the Permian and Triassic periods, was much bigger. It eclipsed both of its predecessors, as well as that which later killed off the dinosaurs—wiping out perhaps 96% of all marine species.
Its after-effects were also much more radical: far from just recovering to former levels, numbers of genera and families eventually grew through the apparent ceiling of the Ordovician to Permian, and continued to do so until the present biodiversity crisis.
How was such a gear change possible? Mass extinctions almost certainly result from catastrophic physical changes to the environment, with a speed that makes it difficult or impossible for animals to adapt and evolve to accommodate. Some groups are depleted much more than others, and in ways that are difficult to predict.

The idea is best illustrated by two groups of clam-like, filter-feeding marine organisms with similar ecologies and life habits: the brachiopods (Phylum Brachiopoda) and the bivalves (Phylum Mollusca). Prior to the end of the Permian, 252 million years ago, brachiopods were much more diverse than bivalves. However, the Great Dying hit the brachiopods much harder than the bivalves, and bivalves also recovered much faster. Not only did the bivalves rise to dominance in the wake of the mass extinction—they went on to become much more diverse than the brachiopods had ever been.
Such a turning of the tables may be possible when one group has already filled an ecospace, making it difficult for other groups to get a foothold. Only rapid change in the physical environment can dislodge them, offering ecological competitors the opportunity they previously lacked. These ascendant groups may also subdivide ecospace more finely (smaller wedges in Darwin’s analogy), allowing a stalled diversity curve to take off again. New species may also change the environment in ways that provide niches for others, thereby creating new ecospace (or enlarging Darwin’s log).
Something of this sort happened on land with the extinction of the dinosaurs at the Cretaceous–Paleogene extinction event some 66 million years ago, which saw mammals comparatively mildly affected. Ironically, the Great Dying event had previously knocked the then hugely successful ancestors of the modern mammals—the therapsids—into the background some 186 million years earlier, allowing the archosaurs and ultimately the dinosaurs to flourish in the first place. What goes around comes around.
With such major shifts in the Earth’s biodiversity seemingly hostage to the whims of fortune, paleontologists have looked for any general rules that might predict survival. On land, large size seems to be disadvantageous.

Alarmingly, few animals larger than a dog survived the Cretaceous–Paleogene event. Other disadvantages include ecological specialization and having a restricted geographical distribution.
In between extinction events, a wide geographic distribution appears to offer considerable insurance. However we have recently shown that geographical range had no effect on the number of surviving terrestrial vertebrate species at the end of the Triassic mass extinction some 201 million years ago. The physical events causing mass extinctions, whether asteroids, mass volcanism or other physical factors, are so disruptive and have such global consequences that even the most widespread and numerous species can be wiped out.
It is, therefore, very difficult to make generalizations and predictions. But we do know that nothing is ever really safe. As we face the prospect of the sixth mass extinction, albeit caused by human activity this time, it is well to remember that extinctions can quickly escalate in unpredictable ways.
The loss of one species can have unforeseen consequences for many others, since ecosystems are connected by a complex web of interactions that we do not always fully understand. We must hope that such an ecosystem collapse is far enough down the road for us to forestall it. Unfortunately, early signs—such as habitat fragmentation and species loss in rainforests and reefs—are not good.